By Alex Liebman and Rob Wallace, The Agroecology and Rural Economics Research Corps, April 10, 2019

St Paul, April 9—Eighty percent of U.S. antibiotics are used to promote livestock and poultry growth and protect the animals from the bacterial consequences of the manure-laden environments in which they are grown. That’s 34 million pounds of antibiotics a year as of 2015.
The agricultural applications help generate drug resistance across multiple human bacterial infections, killing 23,000-100,000 Americans a year and, with an increasing amount of antibiotics applied abroad, 700,000 people worldwide.
Now a fungal species, Candida auris, has developed multidrug resistance and is rapidly spreading across human populations across the globe (see nearby figure). The CDC reports 90% of C. auris infections are clocking in resistant to one antifungal drug and 30% to two or more.
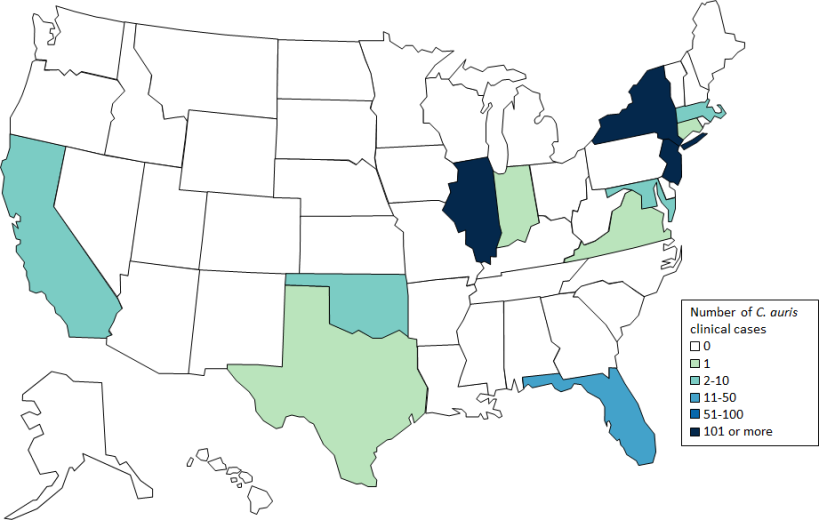
Clinical cases of Candida auris reported by CDC as of February 28, 2019: by country (top) and U.S. state (bottom). From CDC (2019).
C. auris, a yeast, is killing immunocompromised patients in hospitals, clinics, and nursing homes at a prodigious clip, up to 40-60% of those who suffer bloodstream infections in a month’s time.
In the rooms of the infected and the dead, the fungus appears intransigent to nearly all attempts at eradication. The fungus can survive even a floor-to-ceiling spray of aerosolized hydrogen peroxide.
How have drug-resistant fungi come to haunt the modern hospital and jeopardize the sterile spaces asepsis addressed 150 years ago?
It is becoming increasingly apparent that C. auris’s resistance, and that of many other fungi species, is traceable to industrial agriculture’s mass application of fungicides. These chemicals approximate the molecular structures of antifungal drugs.
Across crops—wheat, banana, barley, apple, among many others—the fungicides select for resistant strains that find their way into hospitals where they are also resistant to the drugs administered to patients.
The azoles and morpholines target the ergosterol biosynthetic pathway, which generates the plasma membrane of fungi cells. The benzimidazoles interfere with fungi cytoskeleton, preventing the assembly of cell microtubules. The strobilurins and succinate dehydrogenase inhibitors take more physiological routes, inhibiting the electron transfer chain of mitochondrial respiration. The anilinopyrimidines appear to target mitochondrial signalling pathways.
Candida auris has evolved resistance to a suite of azole antifungals, including fluconazole, with variable susceptibilities to other azoles, amphotericin B, and echinocandins. Azoles, used in both crop protection and medical settings, are broad-spectrum fungicides, annihilating a wide range of fungi rather than targeting a specific type.
How did fungus and fungicide find each other in the field?
C. auris, likely long circulating on its own for thousands of years as CDC’s Tom Chiller hypothesizes, was first isolated in humans from the ear canal of a 70-year-old Japanese woman at a Tokyo hospital in 2009 (although a 1996 isolate was subsequently identified). Later isolation found the yeast capable of bloodstream infection.
In an effort to identify the source of the infection, an international team sequenced resistant isolates collected from hospitals across Pakistan, India, South Africa, and Venezuela, 2012–2015.
Against expectations, the team found divergent amino acid replacements associated with azole resistance among the ERG11 single nucleotide polymorphisms—one among several such SNPs—across four geographic regions. They weren’t the same strain, indicating that each resistant phenotype had emerged independently.
In other words, strains isolated by distance from each other evolved unique solutions to the fungicides to which they were exposed.
That might indicate molecular adaptations to different exposures. But it also might indicate that in response to such wide exposure to fungicides in the field, each strain evolved its own unique solution to the problem.
Even as fungi do not horizontally transfer their genes at rates that virus and bacteria do, migration of patients and fungi alike, the latter by way of agricultural trade, can help increase diversity in the fungicidal resistance circulating in any one locale.
A second team identified multiple genotypes of different international origins in the relatively bounded confines of the United Kingdom. A third team, as the nearby map shows, identified a similar mix in U.S. cases.
But it isn’t clear other than travel-related cases whether all the cases originated from strains from abroad. Without a baseline of fungal load among, say, domestic agricultural workers, an endogenous source remains a possibility.

Distribution of Candida auris clades in the United States. (A) Maximum parsimony phylogenetic tree of marker isolates from Colombia, India, Japan, Pakistan, South Korea, South Africa, Venezuela, and U.S. clinical cases in the USA. (B) The frequency of U.S. clinical cases by clade. (C) The phylogeography of introduced clades. Solid lines indicate introductions that are associated with patients known to have received health care abroad. Adapted from Chow et al. (2018).
To add to the complexity, there also appear multiple mechanisms by which resistance emerges.
Dominique Sanglard summarizes three: decreases in drug concentration in fungal cells, alterations of the drug target, and compensatory mechanisms that depress drug toxicity. Atop these, the three can be arrived at by a variety of genetic events. Alongside SNPs are insertions into the fungus genome, deletions, and structural changes, including gene or chromosome copy events.
One study found 51 genes related to how sensitive circulating strains of a Fusarium blight were to propiconazole, only a single class of triazole fungicide.
The road to such resistance can be complex, winding beyond merely evolving out from underneath a antifungal directly.
In 2015, researchers found that the C. auris genome hosts several genes for the ATP-binding cassette transporter family, a major facilitator superfamily (MFS). MFS transports a large variety of substrates across cell membranes and been shown to effectively dispose of broad classes of drugs. It permits C. auris to survive an onslaught of antifungal drugs.
The team found that that the C. auris genome also encodes a slew of gene families that facilitate the fungi’s virulence. C. auris adaptively forms biofilms that support antifungal resistance by way of a high density of cells, the presence of sterols on biofilm cells, and efficient nutrient use and growth.
One fungus, Aspergillus fumigatus, may offer a conditional preview of C. auris’s trajectories present and future.
Azole antifungals itraconazole, voriconazole, and posaconazole have long been used to treat pulmonary asperillogosis, the infection caused by A. fumigatus. The fungi causes approximately 200,000 deaths per year, in the past decade rapidly developing resistance to antifungal drugs.

Number of peer-reviewed reports of resistance to azole fungicides for plants (in blue) and in humans (in red) for pathogens Aspergillus fumigatus, Candida albicans, C. auris, C. glabrata, Cryptococcus gattii, and Cryptococcus neoformans. From Fisher (2018).
Studies comparing long-term azole users and patients just beginning to take the drug have shown that drug-resistant A. fumigatus was prevalent in both groups, suggesting that resistance evolved in agricultural rather than medical settings.
Researchers have found biogeographical evidence that suggests multi-triazole-resistant A. fumigatus strains in clinical and environmental settings share significant overlap. In the figure nearby, drug resistant A. fumigatus found in the field (green) and in clinical trials (red) map together, demonstrating their coupling in Europe and Asia.

The global map depicts the geographic distribution of multi-triazole-resistant Aspergillus fumigatus strains. Two different mutations are depicted: TR34/L98H (circle) and TR46/Y121F/T289A (square). The percentages denote the environmental prevalence rates of resistance. From Chowdhary et al. (2013).
Other work recently found azole-resistant A. fumigatus related to the use of triazole fungicides in agricultural fields outside of Bogotá, Colombia. Soils were sampled from an array of crop fields and A. fumigatus was grown on agar treated with itraconazole or voriconazole fungicides. In more than 25% of cases, A. fumigatus persisted despite the fungicide treatment.
That is, due to agricultural practices, Aspergillus is entering hospitals already adapted to the slew of antifungal cocktails designed to check its spread. Dumping azoles to control for fungi on grapes, corn, stone fruit, and a myriad of other crops generated the conditions to accelerate drug resistance in human patients.
While extensive phylogenetic and biogeographical research remains to be conducted, a quick perusal of existing distribution maps suggests similarities between Aspergillus fumigatus and its younger (and suddenly more infamous) cohort Candida auris. The strains share similar geographical distributions, occupying many of the same zones described above for C. auris.
The dangers of continuing upon this path of agricultural development are acute.
Medical and agricultural azole fungicides share similar modes of action, so when resistance pops up in one arena it is easily transferable to another. In both agricultural and medical fungicides, the phenyl group of the chemical forms van der Waals contact with the active site of gene cyp51A.
Organic chemistry specifics aside, the close similarities that the Chowdhary group depict in the nearby figure suggest that a mutation in Aspergillus fumigatus to prevent binding to the cyp51A gene in an agricultural setting—specifically a modification of the 14-α sterol demethylase enzyme—would likely confer resistance to medical applications of stereochemically similar drugs.

Diagram showing similar mode of action in triazoles between medical (A) and agricultural (B) applications. From Chowdhary et al. (2013).
Agricultural azole fungicides comprise a third of the total fungicide market. Twenty-five different forms of agricultural azole demethylation inhibitors are in use, compared to just three forms of licensed medical azoles.
So we shouldn’t be surprised that in applying these fungicides at landscape scales in the millions of pounds annually, the medical use of triazole antifungals, using the same mode of action, would rapidly turn ineffective.
Instead of intervening in the interests of global public health to limit these long-problematic applications, government policy in recent years has promoted the lucrative global expansion of fungicide use, fostering the conditions for virulent drug-resistant fungi.
In 2009, fungicides were applied on 30% of corn, soybean, and wheat acreage in the U.S., totaling 80 million acres. Preventative use of fungicides to control soybean rust quadrupled between 2002 and 2006, despite a dubious economic rationale. Global sales continue to skyrocket, nearly tripling since 2005, from $8 billion to $21 billion in 2017.
Fungicides expanded not only in sales but also in geographic distribution.
From the maps nearby, we see tetraconazole, an agricultural triazole, moved from isolated usage in the western Plains in the late 1990s to massive application throughout California’s Central Valley, the upper Midwest, and the Southeast. Boscalid, a fungicide used in fruit and vegetable crops, has increased from ~ 0.15 to 0.6 million pounds from 2004 to 2016, a 400% increase, and is now widely applied across the country.
Estimated agricultural use (EPest-high) of fungicides tetraconazole (left) and boscalid (right) in pounds per U.S. square mile, 1999 and 2014. State-based and other restrictions on pesticide use were not incorporated into EPest-high or EPest-low estimates. EPest-low estimates usually reflect these restrictions because they are based primarily on surveyed data. EPest-high estimates include more extensive estimates of pesticide use not reported in surveys, which sometimes include States or areas when use restrictions have been imposed. Users should consult with State and local agencies for specific use restrictions. National Water-Quality Assessment (NAWQA) Project/USGS/ARERC.
From within each new locale, the fungicides percolate into the local environment.
In 2012, USGS scientists studied 33 different fungicides used in potato production and found at least one fungicide in 75% of tested surface waters and 58% of ground water samples. With half-lives stretching to several months, azole fungicides are able to easily reach and persist in aquatic environments by runoff and spray drift, becoming highly mobile.
As climate change fundamentally reshapes the U.S., bringing higher overall temperatures and extreme oscillations between drought and heavy rainfall, fungi are predicted to expand outside of their current ranges while also responding specifically to new climate regimes. Aspergillus flavus, the producer of a cancer-causing aflatoxin that reduces corn yields and poisons humans, thrives in drought conditions and large crop-water deficits.
With the market treated as a force of nature stronger than climate or public health, under current agricultural production, broad-spectrum fungicide use is likely only to increase.
A 2016 UK report, citing the overapplication of agricultural fungicides, recommended increased surveillance of antibiotic usage overall and a regulatory apparatus organized by the WHO, FAO, and OIE that among its duties would list critical antibiotics that should be barred from agriculture use.
But aside from collecting more information and calling for what appears minimal regulation, what is to be done?
Given recent travails in antibiotic and herbicide resistance, it seems likely that chemical companies and their farming clients will pursue developing new fungicides based on targeted molecular research, multiple drug cocktails, and gene-edited resistance.
Governmental agencies are likely to impose increased if dubious biosecurity measures, which also frequently foment xenophobic anxieties and are used to blame workers for contamination, rather than addressing the systemic failures of industrial agriculture.
The conjoined motives of powerful medical and agricultural companies are almost certain to promote ‘solutions’ that exacerbate an arms race between toxic drug applications and fungal resistance, spew growing permutations of lethal chemicals into the environment, and further consolidate and privatize the agro-pharmaceutical sector.
There is, however, a different, evidence-based paradigm for responding to fungicidal collapse.
A quick review of agroecological examples suggests that a combination of disease modeling and cultural practices such as crop rotation and cover cropping can greatly reduce the presence of fungal diseases and thus dependence on fungicides.

Intercropping, here soybean and flax, can increase and diversify the soil microbiota to exclude pathogenic fungi. Photo: Alexis Stockford.
In the California’s Central Valley, strawberry producers accustomed to fumigating soils with fungicides to control incidence of Verticillium wilt, a pathogenic soil fungi, have found that planting broccoli crops in between rotations of strawberry crops greatly reduced levels of Verticillium.
Dating back several decades, similar results have been found in the diversification of potato crop rotations.
Researchers in India—a country where drug-resistant A. fumigatus and C. auris have both been found—have studied novel approaches to controlling late blight in potato.
Potato crops often receive large doses of azole fungicides to control for fungal pathogens such as late blight. Rather than fungicide treatments, scientists applied silica to foliar tissue, finding that silica was absorbed and strengthened the potato’s cell walls against fungal invasion. Disease infestation rates ranged from 2.8 – 7.9% in the silica-based integrated management systems and 49.4 – 66.7% in the conventional fungicide dependent systems.
In general, organic farming supports mutualistic fungi to a much greater degree than conventional farming, crowding out pathogenic strains. Crop rotations, the incorporation of legumes, and the cultivation of soil aggregates support ecological niches for soil microbiota.
Reducing chemical fertilizers and limiting tillage, two agroecological practices with major benefits for reduced pollution and enhanced carbon storage, also select for beneficial strains of arbuscular mycorrhizal fungi that form mutualistic relationships with plant roots and can confer resistance to soil pathogens.
Integrating agricultural production into a broader matrix of non-crop vegetation is also important for controlling fungal pathogens. Wild landscapes reduce the potential for pathogen populations to adapt to crops and modeling suggests that contiguous swaths of wild patches reduce the aggressiveness of pathogens upon agricultural crops.
Ivette Perfecto and John Vandermeer’s labs have done yeoman work, written up in depth here and summarized here, tracing the means by which thatches of ecological relationships—predation, mutualism, competition, etc.—up and down the food web in which a crop finds itself can box out pest damage, including, their teams find, from rust fungi.

The nitty-gritty as it applies to fungi can be found in Vandermeer student Douglas Jackson’s dissertation on agroecological fungal control in coffee.
Zachary Hajian-Forooshani (pictured), another University of Michigan student, followed up research from the 1970s and found Mycodiplosis fly larvae feed on the coffee rust the Perfecto-Vandermeer team study in Mexico and Puerto Rico.
What emerges is a picture of ecological complexity in which fungicidal warfare is exactly the wrong tool.
Instead, throwing bad money after bad, fungicides today are applied in a system in which diseases thrive out of simplified landscapes, vast and uninterrupted genetically identical monocultures, rapidly accelerating global warming, and an ever quickening pace of global trade.
In a cruel irony, fungicide application places evolutionary pressure on pathogens to develop resistance at the same time that industrial management provides the near-perfect conditions for fostering and spreading these virulent mutations.
It all makes sense only when we recognize that the agribusiness sector views nature as its stiffest competition.
Wiping out local ecologies and the near-free work these offer in helping farmers enrich their soils, clean their water, pollinate their plants, feed their livestock, and control pests—pathogenic fungi among them—means the largest companies can now sell commodified equivalents to a captured market.
The damage done is more than agricultural or economic. It’s a business plan pursued even at the risk of eroding our capacity to socially reproduce ourselves as a civilization.
Farmers and food activists have complained industrial agriculture represents little more than nutrient and carbon mining. Companies are compelling farmers to grow so much so fast that production squeezes carbon out of the soil in the form of food commodities. As a result, land and water are polluted into such oblivion that food safety cannot be accounted for.
By that pollution, occupational exposures, outbreaks of increasing virulence and extent, metabolic diseases such as diabetes, antibiotic resistance, and now the growing threat of fungicide resistance, carbon mining now extends to gouging out global public health.
Once made the order of the day, alternate agricultures long pursued and updated by smallholders worldwide, and backed by a growing scientific literature, offer a way out of that trap.

Alex Liebman is a plant-soil and political ecology researcher with Lurralde, a Chilean group supporting the Atacameña and Ayamara peoples in their struggle for territorial sovereignty and water rights in the face of multinational copper and lithium mining interests in the Atacama Desert.
 Rob Wallace is an evolutionary biologist and public health phylogeographer. He’s the author of Big Farms Make Big Flu and, most recently, co-author of Clear-Cutting Disease Control.
Rob Wallace is an evolutionary biologist and public health phylogeographer. He’s the author of Big Farms Make Big Flu and, most recently, co-author of Clear-Cutting Disease Control.
The ideas and opinions posted here are the authors’ and not necessarily that of ARERC as an organization.
St Paul, April 9—Eighty percent of U.S. antibiotics are used to promote livestock and poultry growth and protect the animals from the bacterial consequences of the manure-laden environments in which they are grown. That’s 34 million pounds of antibiotics a year as of 2015.
The agricultural applications help generate drug resistance across multiple human bacterial infections, killing 23,000-100,000 Americans a year and, with an increasing amount of antibiotics applied abroad, 700,000 people worldwide.
Now a fungal species, Candida auris, has developed multidrug resistance and is rapidly spreading across human populations across the globe (see nearby figure). The CDC reports 90% of C. auris infections are clocking in resistant to one antifungal drug and 30% to two or more.
Clinical cases of Candida auris reported by CDC as of February 28, 2019: by country (top) and U.S. state (bottom). From CDC (2019).
C. auris, a yeast, is killing immunocompromised patients in hospitals, clinics, and nursing homes at a prodigious clip, up to 40-60% of those who suffer bloodstream infections in a month’s time.
In the rooms of the infected and the dead, the fungus appears intransigent to nearly all attempts at eradication. The fungus can survive even a floor-to-ceiling spray of aerosolized hydrogen peroxide.
How have drug-resistant fungi come to haunt the modern hospital and jeopardize the sterile spaces asepsis addressed 150 years ago?
It is becoming increasingly apparent that C. auris’s resistance, and that of many other fungi species, is traceable to industrial agriculture’s mass application of fungicides. These chemicals approximate the molecular structures of antifungal drugs.
Across crops—wheat, banana, barley, apple, among many others—the fungicides select for resistant strains that find their way into hospitals where they are also resistant to the drugs administered to patients.
The path of yeast resistance
Matthew Fisher and colleagues recently classified six main classes of fungicides, all rarely used in the U.S. Midwest before 2007.The azoles and morpholines target the ergosterol biosynthetic pathway, which generates the plasma membrane of fungi cells. The benzimidazoles interfere with fungi cytoskeleton, preventing the assembly of cell microtubules. The strobilurins and succinate dehydrogenase inhibitors take more physiological routes, inhibiting the electron transfer chain of mitochondrial respiration. The anilinopyrimidines appear to target mitochondrial signalling pathways.
Candida auris has evolved resistance to a suite of azole antifungals, including fluconazole, with variable susceptibilities to other azoles, amphotericin B, and echinocandins. Azoles, used in both crop protection and medical settings, are broad-spectrum fungicides, annihilating a wide range of fungi rather than targeting a specific type.
How did fungus and fungicide find each other in the field?
C. auris, likely long circulating on its own for thousands of years as CDC’s Tom Chiller hypothesizes, was first isolated in humans from the ear canal of a 70-year-old Japanese woman at a Tokyo hospital in 2009 (although a 1996 isolate was subsequently identified). Later isolation found the yeast capable of bloodstream infection.
In an effort to identify the source of the infection, an international team sequenced resistant isolates collected from hospitals across Pakistan, India, South Africa, and Venezuela, 2012–2015.
Against expectations, the team found divergent amino acid replacements associated with azole resistance among the ERG11 single nucleotide polymorphisms—one among several such SNPs—across four geographic regions. They weren’t the same strain, indicating that each resistant phenotype had emerged independently.
In other words, strains isolated by distance from each other evolved unique solutions to the fungicides to which they were exposed.
That might indicate molecular adaptations to different exposures. But it also might indicate that in response to such wide exposure to fungicides in the field, each strain evolved its own unique solution to the problem.
Even as fungi do not horizontally transfer their genes at rates that virus and bacteria do, migration of patients and fungi alike, the latter by way of agricultural trade, can help increase diversity in the fungicidal resistance circulating in any one locale.
A second team identified multiple genotypes of different international origins in the relatively bounded confines of the United Kingdom. A third team, as the nearby map shows, identified a similar mix in U.S. cases.
But it isn’t clear other than travel-related cases whether all the cases originated from strains from abroad. Without a baseline of fungal load among, say, domestic agricultural workers, an endogenous source remains a possibility.
Distribution of Candida auris clades in the United States. (A) Maximum parsimony phylogenetic tree of marker isolates from Colombia, India, Japan, Pakistan, South Korea, South Africa, Venezuela, and U.S. clinical cases in the USA. (B) The frequency of U.S. clinical cases by clade. (C) The phylogeography of introduced clades. Solid lines indicate introductions that are associated with patients known to have received health care abroad. Adapted from Chow et al. (2018).
To add to the complexity, there also appear multiple mechanisms by which resistance emerges.
Dominique Sanglard summarizes three: decreases in drug concentration in fungal cells, alterations of the drug target, and compensatory mechanisms that depress drug toxicity. Atop these, the three can be arrived at by a variety of genetic events. Alongside SNPs are insertions into the fungus genome, deletions, and structural changes, including gene or chromosome copy events.
One study found 51 genes related to how sensitive circulating strains of a Fusarium blight were to propiconazole, only a single class of triazole fungicide.
The road to such resistance can be complex, winding beyond merely evolving out from underneath a antifungal directly.
In 2015, researchers found that the C. auris genome hosts several genes for the ATP-binding cassette transporter family, a major facilitator superfamily (MFS). MFS transports a large variety of substrates across cell membranes and been shown to effectively dispose of broad classes of drugs. It permits C. auris to survive an onslaught of antifungal drugs.
The team found that that the C. auris genome also encodes a slew of gene families that facilitate the fungi’s virulence. C. auris adaptively forms biofilms that support antifungal resistance by way of a high density of cells, the presence of sterols on biofilm cells, and efficient nutrient use and growth.
Other fungi, other dangers
Candida auris is hardly the only deadly fungus converging upon multidrug resistance. The nearby map shows multiple species overlapping in plant and human resistance.One fungus, Aspergillus fumigatus, may offer a conditional preview of C. auris’s trajectories present and future.
Azole antifungals itraconazole, voriconazole, and posaconazole have long been used to treat pulmonary asperillogosis, the infection caused by A. fumigatus. The fungi causes approximately 200,000 deaths per year, in the past decade rapidly developing resistance to antifungal drugs.
Number of peer-reviewed reports of resistance to azole fungicides for plants (in blue) and in humans (in red) for pathogens Aspergillus fumigatus, Candida albicans, C. auris, C. glabrata, Cryptococcus gattii, and Cryptococcus neoformans. From Fisher (2018).
Studies comparing long-term azole users and patients just beginning to take the drug have shown that drug-resistant A. fumigatus was prevalent in both groups, suggesting that resistance evolved in agricultural rather than medical settings.
Researchers have found biogeographical evidence that suggests multi-triazole-resistant A. fumigatus strains in clinical and environmental settings share significant overlap. In the figure nearby, drug resistant A. fumigatus found in the field (green) and in clinical trials (red) map together, demonstrating their coupling in Europe and Asia.
The global map depicts the geographic distribution of multi-triazole-resistant Aspergillus fumigatus strains. Two different mutations are depicted: TR34/L98H (circle) and TR46/Y121F/T289A (square). The percentages denote the environmental prevalence rates of resistance. From Chowdhary et al. (2013).
Other work recently found azole-resistant A. fumigatus related to the use of triazole fungicides in agricultural fields outside of Bogotá, Colombia. Soils were sampled from an array of crop fields and A. fumigatus was grown on agar treated with itraconazole or voriconazole fungicides. In more than 25% of cases, A. fumigatus persisted despite the fungicide treatment.
That is, due to agricultural practices, Aspergillus is entering hospitals already adapted to the slew of antifungal cocktails designed to check its spread. Dumping azoles to control for fungi on grapes, corn, stone fruit, and a myriad of other crops generated the conditions to accelerate drug resistance in human patients.
While extensive phylogenetic and biogeographical research remains to be conducted, a quick perusal of existing distribution maps suggests similarities between Aspergillus fumigatus and its younger (and suddenly more infamous) cohort Candida auris. The strains share similar geographical distributions, occupying many of the same zones described above for C. auris.
Industrial agriculture’s role
With zones of overlapping human and crop resistant cases of Aspergillus fumigatus and the rising specter of a new azole resistant fungus ravaging clinical settings and evolving at lightning speed, one would hope that azole fungicide use would be closely monitored if not just phased out.The dangers of continuing upon this path of agricultural development are acute.
Medical and agricultural azole fungicides share similar modes of action, so when resistance pops up in one arena it is easily transferable to another. In both agricultural and medical fungicides, the phenyl group of the chemical forms van der Waals contact with the active site of gene cyp51A.
Organic chemistry specifics aside, the close similarities that the Chowdhary group depict in the nearby figure suggest that a mutation in Aspergillus fumigatus to prevent binding to the cyp51A gene in an agricultural setting—specifically a modification of the 14-α sterol demethylase enzyme—would likely confer resistance to medical applications of stereochemically similar drugs.
Diagram showing similar mode of action in triazoles between medical (A) and agricultural (B) applications. From Chowdhary et al. (2013).
Agricultural azole fungicides comprise a third of the total fungicide market. Twenty-five different forms of agricultural azole demethylation inhibitors are in use, compared to just three forms of licensed medical azoles.
So we shouldn’t be surprised that in applying these fungicides at landscape scales in the millions of pounds annually, the medical use of triazole antifungals, using the same mode of action, would rapidly turn ineffective.
Instead of intervening in the interests of global public health to limit these long-problematic applications, government policy in recent years has promoted the lucrative global expansion of fungicide use, fostering the conditions for virulent drug-resistant fungi.
In 2009, fungicides were applied on 30% of corn, soybean, and wheat acreage in the U.S., totaling 80 million acres. Preventative use of fungicides to control soybean rust quadrupled between 2002 and 2006, despite a dubious economic rationale. Global sales continue to skyrocket, nearly tripling since 2005, from $8 billion to $21 billion in 2017.
Fungicides expanded not only in sales but also in geographic distribution.
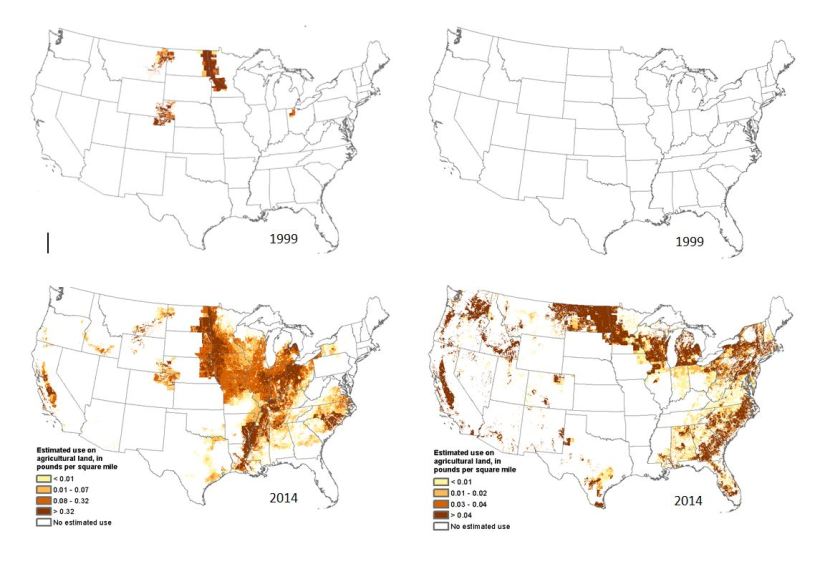
From the maps nearby, we see tetraconazole, an agricultural triazole, moved from isolated usage in the western Plains in the late 1990s to massive application throughout California’s Central Valley, the upper Midwest, and the Southeast. Boscalid, a fungicide used in fruit and vegetable crops, has increased from ~ 0.15 to 0.6 million pounds from 2004 to 2016, a 400% increase, and is now widely applied across the country.
Estimated agricultural use (EPest-high) of fungicides tetraconazole (left) and boscalid (right) in pounds per U.S. square mile, 1999 and 2014. State-based and other restrictions on pesticide use were not incorporated into EPest-high or EPest-low estimates. EPest-low estimates usually reflect these restrictions because they are based primarily on surveyed data. EPest-high estimates include more extensive estimates of pesticide use not reported in surveys, which sometimes include States or areas when use restrictions have been imposed. Users should consult with State and local agencies for specific use restrictions. National Water-Quality Assessment (NAWQA) Project/USGS/ARERC.
From within each new locale, the fungicides percolate into the local environment.
In 2012, USGS scientists studied 33 different fungicides used in potato production and found at least one fungicide in 75% of tested surface waters and 58% of ground water samples. With half-lives stretching to several months, azole fungicides are able to easily reach and persist in aquatic environments by runoff and spray drift, becoming highly mobile.
As climate change fundamentally reshapes the U.S., bringing higher overall temperatures and extreme oscillations between drought and heavy rainfall, fungi are predicted to expand outside of their current ranges while also responding specifically to new climate regimes. Aspergillus flavus, the producer of a cancer-causing aflatoxin that reduces corn yields and poisons humans, thrives in drought conditions and large crop-water deficits.
With the market treated as a force of nature stronger than climate or public health, under current agricultural production, broad-spectrum fungicide use is likely only to increase.
Farming as its own fungus control
In response to drug-resistant bacteria and fungi, research institutions are calling for the collection of better data on agricultural antibiotic use and on the potential economic costs of transitioning away from from high rates of application.A 2016 UK report, citing the overapplication of agricultural fungicides, recommended increased surveillance of antibiotic usage overall and a regulatory apparatus organized by the WHO, FAO, and OIE that among its duties would list critical antibiotics that should be barred from agriculture use.
But aside from collecting more information and calling for what appears minimal regulation, what is to be done?
Given recent travails in antibiotic and herbicide resistance, it seems likely that chemical companies and their farming clients will pursue developing new fungicides based on targeted molecular research, multiple drug cocktails, and gene-edited resistance.
Governmental agencies are likely to impose increased if dubious biosecurity measures, which also frequently foment xenophobic anxieties and are used to blame workers for contamination, rather than addressing the systemic failures of industrial agriculture.
The conjoined motives of powerful medical and agricultural companies are almost certain to promote ‘solutions’ that exacerbate an arms race between toxic drug applications and fungal resistance, spew growing permutations of lethal chemicals into the environment, and further consolidate and privatize the agro-pharmaceutical sector.
A quick review of agroecological examples suggests that a combination of disease modeling and cultural practices such as crop rotation and cover cropping can greatly reduce the presence of fungal diseases and thus dependence on fungicides.
Intercropping, here soybean and flax, can increase and diversify the soil microbiota to exclude pathogenic fungi. Photo: Alexis Stockford.
In the California’s Central Valley, strawberry producers accustomed to fumigating soils with fungicides to control incidence of Verticillium wilt, a pathogenic soil fungi, have found that planting broccoli crops in between rotations of strawberry crops greatly reduced levels of Verticillium.
Dating back several decades, similar results have been found in the diversification of potato crop rotations.
Researchers in India—a country where drug-resistant A. fumigatus and C. auris have both been found—have studied novel approaches to controlling late blight in potato.
Potato crops often receive large doses of azole fungicides to control for fungal pathogens such as late blight. Rather than fungicide treatments, scientists applied silica to foliar tissue, finding that silica was absorbed and strengthened the potato’s cell walls against fungal invasion. Disease infestation rates ranged from 2.8 – 7.9% in the silica-based integrated management systems and 49.4 – 66.7% in the conventional fungicide dependent systems.
In general, organic farming supports mutualistic fungi to a much greater degree than conventional farming, crowding out pathogenic strains. Crop rotations, the incorporation of legumes, and the cultivation of soil aggregates support ecological niches for soil microbiota.
Reducing chemical fertilizers and limiting tillage, two agroecological practices with major benefits for reduced pollution and enhanced carbon storage, also select for beneficial strains of arbuscular mycorrhizal fungi that form mutualistic relationships with plant roots and can confer resistance to soil pathogens.
Integrating agricultural production into a broader matrix of non-crop vegetation is also important for controlling fungal pathogens. Wild landscapes reduce the potential for pathogen populations to adapt to crops and modeling suggests that contiguous swaths of wild patches reduce the aggressiveness of pathogens upon agricultural crops.
Ivette Perfecto and John Vandermeer’s labs have done yeoman work, written up in depth here and summarized here, tracing the means by which thatches of ecological relationships—predation, mutualism, competition, etc.—up and down the food web in which a crop finds itself can box out pest damage, including, their teams find, from rust fungi.
The nitty-gritty as it applies to fungi can be found in Vandermeer student Douglas Jackson’s dissertation on agroecological fungal control in coffee.
Zachary Hajian-Forooshani (pictured), another University of Michigan student, followed up research from the 1970s and found Mycodiplosis fly larvae feed on the coffee rust the Perfecto-Vandermeer team study in Mexico and Puerto Rico.
More than mining soil
All this work squares well with agroecological theory that under current political policies and demographic trends, farm fields integrated into a matrix of nature conservation are more likely than ‘land-sparing’ approaches to conserve natural resources while simultanWhat emerges is a picture of ecological complexity in which fungicidal warfare is exactly the wrong tool.
Instead, throwing bad money after bad, fungicides today are applied in a system in which diseases thrive out of simplified landscapes, vast and uninterrupted genetically identical monocultures, rapidly accelerating global warming, and an ever quickening pace of global trade.
In a cruel irony, fungicide application places evolutionary pressure on pathogens to develop resistance at the same time that industrial management provides the near-perfect conditions for fostering and spreading these virulent mutations.
It all makes sense only when we recognize that the agribusiness sector views nature as its stiffest competition.
Wiping out local ecologies and the near-free work these offer in helping farmers enrich their soils, clean their water, pollinate their plants, feed their livestock, and control pests—pathogenic fungi among them—means the largest companies can now sell commodified equivalents to a captured market.
The damage done is more than agricultural or economic. It’s a business plan pursued even at the risk of eroding our capacity to socially reproduce ourselves as a civilization.
Farmers and food activists have complained industrial agriculture represents little more than nutrient and carbon mining. Companies are compelling farmers to grow so much so fast that production squeezes carbon out of the soil in the form of food commodities. As a result, land and water are polluted into such oblivion that food safety cannot be accounted for.
By that pollution, occupational exposures, outbreaks of increasing virulence and extent, metabolic diseases such as diabetes, antibiotic resistance, and now the growing threat of fungicide resistance, carbon mining now extends to gouging out global public health.
Once made the order of the day, alternate agricultures long pursued and updated by smallholders worldwide, and backed by a growing scientific literature, offer a way out of that trap.
Alex Liebman is a plant-soil and political ecology researcher with Lurralde, a Chilean group supporting the Atacameña and Ayamara peoples in their struggle for territorial sovereignty and water rights in the face of multinational copper and lithium mining interests in the Atacama Desert.
Rob Wallace is an evolutionary biologist and public health phylogeographer. He’s the author of Big Farms Make Big Flu and, most recently, co-author of Clear-Cutting Disease Control.The ideas and opinions posted here are the authors’ and not necessarily that of ARERC as an organization.
No comments:
Post a Comment